
Page 149 - 《广西植物》2022年第8期
P. 149
ϟ ሕ ँ ऐ 广জ 西জ 植জ 物 ࿗ 卷
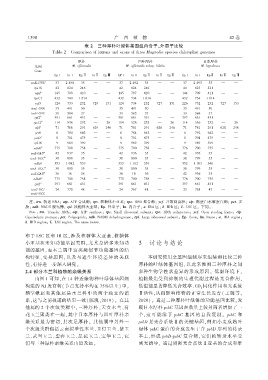
表 ԣই 三种厚朴叶绿体基因组内含子ܓ外显子比较
ఋᑕᤦᔀ জ ऊᆍቝᡱᑕʢᔽ༁ᆍႿ ᆍ ᔽႿᡷʢᆍႿ༁ ᑕႿʛ ᔀᆍႿ༁ ᆍ ᡷၤʢᔀᔀ ᤹ᐹႥᅳᤀᔠᐹ ༁ᡱᔀዹᔽᔀ༁ ዹၤᆍʢᆍᡱᑕ༁ᡷ ੫ᔀႿᆍቝᔀ༁
厚朴 凹叶厚朴 日本厚朴
基因 ᤹ॹ ᅳஏஏᔠዞᔠႥᐹᤀᔠ ᤹ॹ ᅳஏஏᔠዞᔠႥᐹᤀᔠ ༁ࣼᤦ༁ᡱ ᤇᔠᤀᅳᤇᐹ ᤹ॹ ၊ᡥᡓᅳᤀᓣࣩዞᐹ
᥈ᔀႿᔀ
ጶᡱ ᧧ ᧧Ⴟ ᧧ ጶᡱ֒ ᧧Ⴟ֒ ጶᡱᎏ ጶ۪ ᧧ ᧧Ⴟ ᧧ ጶᡱ֒ ᧧Ⴟ֒ ጶᡱᎏ ጶᡱ ᧧ ᧧Ⴟ ᧧ ጶᡱ֒ ᧧Ⴟ֒ ጶᡱᎏ
ᡙʗႥᣞІІІ ሕᎮ ࿗ँऐ ሕጢ ᗵ ᗵ ሕᎮ ࿗ँ ሕጢ ᗵ ᗵ ሕᎮ ࿗ँሕ ሕጢ ᗵ ᗵ
ʗᡓϟᤃ ࿗ ऐ࿗ ࿗ᤃ ᗵ ᗵ ࿗ ऐ࿗ ࿗ᤃ ᗵ ᗵ ࿗ԡ ऐሕ ϟ ᗵ ᗵ
ᐹᡙᡓÞ ϟ࿗ጢ Ꭾԡँ ࿗ϟԡ ᗵ ᗵ ϟ࿗ጢ ᎮԡᎮ ࿗ϟԡ ᗵ ᗵ ϟ࿗࿗ Ꭾԡँ ࿗ϟϟ ᗵ ᗵ
ʗᡓᅳࣷϟ ࿗ሕ Ꭾ࿗ԡ ϟ ᤃϟ࿗ ᗵ ᗵ ࿗ሕ Ꭾሕ࿗ ϟ ᤃϟ࿗ ᗵ ᗵ ࿗ሕ Ꭾሕ࿗ ϟ ᤃϟ࿗ ᗵ ᗵ
ᡥዞஏሕ ϟ࿗ Ꭾሕሕ ሕ Ꭾँ ϟጢϟ ϟ࿗ Ꭾሕ࿗ ሕ ᎮᎮ ϟጢϟ ᤃ Ꭾሕ ሕ ᎮᎮ ϟጢሕ
ᡙʗႥᒦІዛዛ ሕጢ ࿗ँϟ ጢԡ ᗵ ᗵ ሕጢ ࿗ँϟ ጢԡ ᗵ ᗵ ሕጢ ࿗ँϟ ጢԡ ᗵ ᗵ
ᡙʗႥႬІዛࣷ ሕँ ጢऐ࿗ ሕᎮ ሕँ ጢऐጢ ሕᎮ ሕँ ጢऐ࿗ ሕᎮ
ᤇ ᗵ ᗵ
ʗᡓᤀ ሕँϟ ᤃᤃϟ ࿗ሕϟ ᗵ ᗵ ሕँϟ ᤃᤃϟ ሕँϟ ᗵ ᗵ ሕँᎮ ᤃᤃϟ ࿗ሕϟ
ᤇ
ʗᡓϟ ϟϟ࿗ ጢሕᤃ ሕ ᗵ ᤃ ϟϟ࿗ ጢᤃ ሕ ᗵ ᤃ ϟϟ࿗ ጢሕᤃ ሕ ᗵ ᤃ
ዞᤀᡓۗ Ꭾϟ Ꭾऐᤃ ँϟ ᤃँ ࿗ᤃ Ꭾϟ Ꭾऐϟ ँϟ ᤃऐ ࿗ᤃ Ꭾϟ Ꭾऐϟ ँϟ ᤃऐ ࿗ᤃ
ᡓᓣᡙଧ ᤃ Ꭾऐ࿗ ᤃ࿗ ᗵ ᗵ ᤃ Ꭾऐ࿗ ᤃ࿗ ᗵ ᗵ ᤃ Ꭾँ ᤃ࿗ ᗵ ᗵ
ᡓᓣᡙᦵ ᒦ ऐ Ꭾԡϟ ࿗Ꭾጢ ᗵ ᗵ ऐ Ꭾԡϟ ࿗Ꭾጢ ᗵ ᗵ ऐ Ꭾԡϟ ࿗Ꭾጢ ᗵ ᗵ
ʗᡓᤀϟᤃ ँ ँᤃँ ሕँँ ᗵ ᗵ ँ ँᤃँ ሕँँ ᗵ ᗵ ँ ँᤃँ ሕँँ ᗵ ᗵ
Ⴅʐ၊ଧ ᤇ ᎮᎮጢ Ꭾԡԡ Ꭾጢऐ ᗵ ᗵ ᎮᎮጢ Ꭾԡԡ Ꭾጢऐ ᗵ ᗵ ᎮᎮᤃ Ꭾԡԡ Ꭾጢጢ ᗵ ᗵ
ᡙʗႥᧇᤩዛІ ᐹ ࿗ ँሕᎮ ሕጢ ᗵ ᗵ ࿗ ँሕᤃ ሕጢ ᗵ ᗵ ࿗ ँሕᤃ ሕጢ ᗵ ᗵ
ᡙʗႥዛІᤩࣷ ᤇ ሕऐ ऐԡԡ ሕጢ ᗵ ᗵ ሕऐ ऐԡԡ ሕጢ ᗵ ᗵ ሕऐ Ꭾँँ ሕጢ ᗵ ᗵ
Ⴅʐ၊ዛ ጢጢሕ ϟ ԡऐ ጢሕँ ᗵ ᗵ ጢጢሕ ϟ ϟԡ ጢሕँ ᗵ ᗵ ጢጢ ϟ ϟԡሕ ጢ࿗ԡ ᗵ ᗵ
ᡙʗႥዛІᤩࣷ ᐹ ሕऐ ऐԡԡ ሕጢ ᗵ ᗵ ሕऐ ऐԡԡ ሕጢ ᗵ ᗵ ሕऐ Ꭾँँ ሕጢ ᗵ ᗵ
ᡙʗႥᧇᤩዛІ ᤇ ሕᤃ ϟᤃ ሕᤃ ᗵ ᗵ ሕᤃ ϟᤃ ሕᤃ ᗵ ᗵ ࿗ ँሕᤃ ሕጢ ᗵ ᗵ
Ⴅʐ၊ଧ ᐹ ᎮᎮጢ Ꭾԡԡ Ꭾጢऐ ᗵ ᗵ ᎮᎮጢ Ꭾԡԡ Ꭾጢऐ ᗵ ᗵ ᎮᎮᤃ Ꭾԡԡ Ꭾጢጢ ᗵ ᗵ
ᐹ
ʗᡓᤀ ሕँϟ ᤃᤃϟ ࿗ሕϟ ᗵ ᗵ ሕँϟ ᤃᤃϟ ࿗ሕϟ ᗵ ᗵ ሕँᎮ ᤃᤃϟ ࿗ሕϟ ᗵ ᗵ
ᡙʗႥᤩІࣷ ᣑ ࿗ ᎮᎮԡ ࿗ऐ ᗵ ᗵ ࿗ ᎮᤃᎮ ࿗ऐ ᗵ ᗵ ሕ Ꭾᤃऐ ࿗Ꭾ ᗵ ᗵ
ᡙʗႥᤩᤩࣷࣷ
জ 注᧥ ᡙʗႥ 转运 શዶଫ ᐹᡙᡓዶఋ۪ 合成酶ଫ ʗᡓ 核糖体小亚基ଫ ʗᡓᅳ શዶ 聚合酶ଫ ᡥዞஏ 开放阅读框ଫ ዞᤀᡓ 酪蛋白水解蛋白酶ଫ ᡓᓣᡙ 多
肽ଫ Ⴅʐ၊ શዶ᧕̀ 脱氢酶ଫ ʗᡓᤀ 核糖体大亚基ଫ ጛᡓ 外显子ଫ ᧇႥ 内含子ଫ ᐹ᧧ዶ 区ଫ ᤇ᧧ା 区ଫ ᒦᓂ୩ऊ 区ᤥ 下同ᤥ
জશᆍᡷᔀ᧥ ᡙʗႥ ఋʢᑕႿ༁ᔀʢ શዶଫ ᐹᡙᡓ ዶఋ۪ ༁ᢃႿᡷၤᑕ༁ᔀଫ ʗᡓ ୩ቝᑕ ʢᔽᤦᆍ༁ᆍቝᑕ ༁ࣼᤦࣼႿᔽᡷଫ ʗᡓᅳ શዶ ᡱᆍᢃቝᔀʢᑕ༁ᔀଫ ᡥዞஏ ᨃᡱᔀႿ ʢᔀᑕʛᔽႿ੫ ʢᑕቝᔀଫ ዞᤀᡓ
ऊᑕ༁ᔀᔽႿᆍᢃᡷᔽዹ ᡱʢᆍᡷᔀᑕ༁ᔀଫ ᡓᓣᡙ ۪ᆍᢃᡱᔀᡱᡷᔽʛᔀଫ Ⴅʐ၊ શዶ᧕̀ ʛᔀၤᢃʛʢᆍ੫ᔀႿᑕ༁ᔀଫ ʗᡓᤀ ᓂᑕʢ੫ᔀ ʢᔽᤦᆍ༁ᆍቝᑕ ༁ࣼᤦࣼႿᔽᡷଫ ጛᡓጶᆍႿଫ ᧇႥ᧧ႿᡷʢᆍႿଫ ᐹ᧧ዶ ʢᔀ੫ᔽᆍႿଫ
ᤇ ᧧ା ʢᔀ੫ᔽᆍႿଫ ᒦ ᓂ୩ऊ ʢᔀ੫ᔽᆍႿ ఋၤᔀ ༁ᑕቝᔀ ᤦᔀᆍऔ
布于 ᓂ୩ऊ 区和 ᧧ 区ᖔ涉及核糖体大亚基ܦ核糖体
小亚基和未知功能基因类群ᖔ尤其是诸多未知功 ሕজ 讨论与结论
能的基因ᖔ而木兰属中这类疑似非功能基因的结
构特征ܦ变异原因ᖔ以及与适生环境差异的关联 本研究使用全基因组测序来组装和比较三种
性ᖔ有待进一步深入研究ᤥ 厚朴的叶绿体基因组ᖔ以此来推测三种厚朴之间
ԣॹ 部分木兰科植物的亲缘关系 多种生物学性状差异的形成原因ᤥ 低温环境下ᖔ
由图 ϟ 可知ᖔ在 ϟ࿗ 种近缘物种叶绿体基因组 植物最先受到抑制的生理代谢过程是光合作用ᖔ
构建的 શȮ 发育树ྉ节点支持率均在 Ꭾጢᠮ以上ɯ 中ᖔ 低温能显著降低光合效率ܦऊᨃ 同化作用和光系统
鹅掌楸属和其他属是木兰科中的两个独立的谱 ֒活性ᖔ从而影响植物的正常生长发育ྉ 王璐等ᖔ
系ᖔ这与之前报道的结果一致ྉ 陈凯ᖔԡϟँɯᤥ 在其 ԡԡɯᤥ 通过三种厚朴叶绿体的功能基因比较ᖔ发
他属的 个次级类群中ᖔ三种厚朴ܦ天女木兰ܦ荷 现日本厚朴 ᡓᤇࣷ 基因在数量上较另两者增加了一
花玉兰聚类在一起ᖔ其中日本厚朴与凹叶厚朴亲 个ᖔ这可能缘于 ᡓᤇࣷ 基因的自我复 制ᤥ ᡓᤇࣷ 和
缘关系最为密切ᖔ其次是厚朴ᤥ 其他属中另外一 ᡓᤇᦵ 是光合系统֒的关键基因ᖔ例如小麦成熟叶
个次级类群包括云南拟单性木兰ܦ玉灯玉兰ܦ紫玉 绿体 ᡱ༁ᤦऊ 蛋白的合成发生于含 ᡓᤇᦵ 序列的转录
兰ܦ武当玉兰ܦ望春玉兰ܦ星花玉兰ܦ宝华玉兰ᖔ它 本上ᖔ形成 ᡓᤇᦵ᥋ᡓᤇࣷ 复合物ᖔ它们的转录水平受
们与三种厚朴亲缘关系由近及远ᤥ 到光诱导ᖔ 通过增强光合系统֒亚基的合成和维