
Page 33 - 《广西植物》2020年第2期
P. 33
1 7 0 广 西 植 物 40 卷
Shannon ̄Wiener 指数为 1.203 6 ~ 1.273 5ꎮ 贺治州
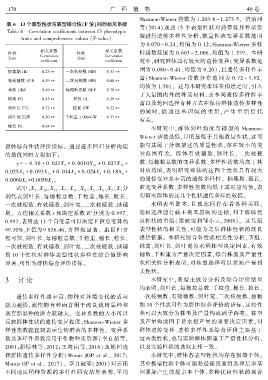
表 6 13 个表型性状与表型综合值(F 值)间的相关系数
等(2014)通过 15 个表型性状对热带优异种质资
Table 6 Correlation coefficients between 13 phenotypic
源进行遗传多样性分析ꎬ数量性状变异系数范围
traits and comprehensive values (F ̄value)
为 0.070 ~ 0.24ꎬ均值为 0.12ꎬShannon ̄Wiener 多样
相关系数 相关系数
性状 性状 性指数范围为 0.603 ~ 2.066ꎬ均值为 1.579ꎮ 本研
Correlation Correlation
Trait Trait
coefficient coefficient 究中ꎬ研究群体具有较大的遗传变异( 变异系数范
围为 0.080 ~ 0.41、均值为 0.20)ꎬ且遗传多样性丰
抽穗期 HD 0.23∗∗ 一次枝梗数 PBN 0.33∗∗
富(Shannon ̄Wiener 指数分布范围为 0.72 ~ 1.92、
有效穗数 EPN 0.19∗∗ 二次枝梗数 SBN 0.68∗∗
均值为 1.50)ꎮ 这与本研究群体在构建之时ꎬ引入
茎粗 DMS 0.46∗∗ 每穗粒总数 NPP 0.70∗∗
了大量国内外的种质材料ꎬ亲本间遗传多样性丰
株高 PH 0.45∗∗ 粒长 GL 0.29∗∗
富以及轮回选择育种方式在保持群体遗传多样性
剑叶长 FLL 0.72∗∗ 粒宽 GW 0.22∗∗
的同 时ꎬ 能 通 过 基 因 间 的 重 组ꎬ 产 生 重 组 后 代
剑叶宽 FLW 0.50∗∗ 千粒重 1 000 ̄GW 0.37∗∗
有关ꎮ
穗长 PL 0.64∗∗
本研究中ꎬ群体剑叶角度与穗型的 Shannon ̄
Wiener 指数最低ꎬ且明显低于其他数量性状ꎬ这可
群体综合性状评价指标ꎮ 通过逐步回归分析构建 能与其属于分级描述的质量性状ꎬ存在较小的变
异范围有关ꎮ 群体有效穗数、剑叶长、二次枝梗
的最优回归方程如下:
y = - 4. 56 + 0. 021X + 0. 0010X + 0. 027X + 数、每穗粒总数的变异系数、多样性指数均高于其
5 10 13
对应均值ꎬ表明研究群体的这四个性状具有较大
0.025X + 0. 093X + 0. 014X + 0. 024X + 0. 18X +
7 11 8 2 6
0.0060X +0.0050X ꎮ 的遗传变异及丰富的遗传多样性ꎮ 抽穗期、穗长、
9 1
式中ꎬX 、X 、X 、X 、X 、X 、X 、X 、X 、X 分 粒宽变异系数、多样性指数均低于其对应均值ꎬ表
5 10 13 7 11 8 2 6 9 1
别代表剑叶长、每穗粒总数、千粒重、穗长、粒长、 明研究群体的这几个性状遗传多样性较低ꎮ
一次枝梗数、有效穗数、剑叶宽、二次枝梗数、抽穗 水稻表 型 繁 多ꎬ且 彼 此 间 存 在 着 各 种 关 联ꎮ
2
期ꎮ 方程相关系数 r 和决定系数 R 分别为 0.997、 轮回选择能打破不利基因间的连锁ꎬ利于群体优
0.993ꎬ表明这 11 个自变量可以决定 F 值总变异的 良性状的重组( 漆映雪和邹小云ꎬ2008)ꎮ 这几组
99.30%ꎬF 值为9 826.46ꎬ方程极显著ꎮ 由回归方 表型性状的相关性ꎬ可能为之后群体性状的改良
程可知ꎬ剑叶长、每穗粒总数、千粒重、穗长、粒长、 提供依据ꎮ 本研究综合各性状相关性分析ꎬ茎粗、
一次枝梗数、有效穗数、剑叶宽、二次枝梗数、抽穗 株高、剑叶长、剑叶宽为水稻株型决定因素ꎬ有效
期 10 个性状对群体表型性状多样性综合值影响 穗数、千粒重为产量决定因素ꎬ综合株型及产量性
显著ꎬ可作为群体综合评价指标ꎮ 状相关性分析表明ꎬ对株型选择可以兼顾产量相
关性状ꎮ
3 讨论 本研究中ꎬ表型主成分分析及综合评价结果
均表明ꎬ剑叶长、每穗粒总数、千粒重、穗长、粒长、
遗传多样性越丰富ꎬ物种对环境变化的适应 一次枝梗数、有效穗数、剑叶宽、二次枝梗数、抽穗
能力越强ꎬ就作物育种而言用于改良栽培品种和 期 10 个性状可作为群体综合评价的指标ꎬ这些性
选育新品种的潜力就越大ꎮ 变异系数的大小可以 状可以大致分为株型及产量构成因子两类ꎮ 株型
反映群体性状的遗传变异程度ꎬShannon ̄Wiener 多 及产量构成因子是水稻产量的重要决定因素ꎬ对
样性指数能直观表示生物群落的多样性ꎮ 变异系 群体遗传变异、遗传多样性及综合评价主要基于
数及多样性指数常用于作物种质资源( 李自超等ꎬ 这两类性状ꎬ也与实验群体侧重于产量性状分析ꎬ
2001ꎻ胡标林等ꎬ2012ꎻ王海岗等ꎬ2016) 及轮回选 以及实验性状的选取上相一致ꎮ
择群体遗传多样性分析( Morais JOP et al.ꎬ 2017ꎻ 本研究中ꎬ群体各表型性状均存在极端个体ꎬ
Morais OP et al.ꎬ 2017)ꎮ 李自超等(2001) 对云南 其中极端性状个体可能是经轮回重组选择后多基
不同地区稻种资源的多样性研究结果表明ꎬ平均 因累加产生的超亲本个体ꎬ多种优良性状的聚合