
Page 125 - 广西植物2024年1期
P. 125
1 期 许爱祝等: 三种地宝兰属植物叶片解剖结构及光合特性的比较研究 1 2 1
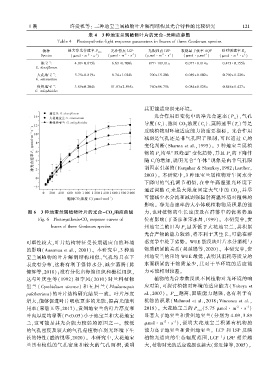
表 4 3 种地宝兰属植物叶片的光合-光响应参数
Table 4 Photosynthetic ̄light response parameters in leaves of three Geodorum species
物种 最大净光合速率 P max 光补偿点 LCP 光饱和点 LSP 表观量子效率 AQY 暗呼吸速率 R d
 ̄2
 ̄1
 ̄1
 ̄1
 ̄2
 ̄2
 ̄2
 ̄1
 ̄1
Species (μmolm s ) (μmolm s ) (μmolm s ) (μmolμmol ) (μmolm s )
地宝兰 4.09±0.675b 6.63±0.708b 877±109.01a 0.077±0.014a 0.473±0.155b
G. densiflorum
大花地宝兰 5.75±0.819a 8.74±1.024b 700±15.20b 0.089±0.002a 0.709±0.226a
G. attenuatum
贵州地宝兰 3.89±0.204b 11.83±2.595a 760±86.75b 0.084±0.028a 0.846±0.427a
G. eulophioides
其更能适应弱光环境ꎮ
光合作用日变化中的净光合速率( P )、气孔
n
导度(G )、胞间 CO 浓度(C )、蒸腾速率(T ) 等是
r
s
i
2
反映植物对环境适应能力的重要指标ꎮ 光合作用
减弱是气孔还是非气孔因子限制ꎬ可以通过 C 的
i
变化判断( Sharma et al.ꎬ1995)ꎮ 3 种地宝兰属植
物的 P 均呈“双峰型”变化趋势ꎬ并且 P 的下降伴
n
n
随 C 的增加ꎬ说明光合“ 午休” 现象是由非气孔限
i
制因素引起的(Farquhar & Shatrkeyꎬ1982ꎻLarcherꎬ
2003)ꎮ 本研究中ꎬ3 种地宝兰属植物对午间水分
下降时的气孔调节相似ꎬ在中午高温强光环境下
通过调整 G 来最大限度固定大气中的 CO ꎬ并尽
s 2
可能减少水分流和减弱强辐射高温环境对植株的
影响ꎮ 净光合速率的大小体现植物物质积累的能
图 6 3 种地宝兰属植物叶片的光合-CO 响应曲线 力ꎬ也对植物的生长速度及在群落中的优劣势地
2
Fig. 6 Photosynthesis ̄CO response curves of 位有影响(丁圣彦和宋永昌ꎬ1999)ꎮ 本研究中ꎬ贵
2
leaves of three Geodorum species 州地宝兰的日均 P 显著低于大花地宝兰ꎬ其积累
n
光合产物的能力较弱ꎬ将不利于其生长ꎬ可能在群
可塑性较大ꎬ叶片结构特征受长期适应自然环境 落竞争中处于劣势ꎮ WUE 能反映叶片水分消耗与
的影响(Aasamaa et al.ꎬ 2001)ꎮ 本研究中ꎬ3 种地 物质积累的关系(吴延娟等ꎬ2020)ꎮ 本研究中ꎬ贵
宝兰属植物的叶片解剖结构相似ꎬ气孔均只在下 州地宝兰的日均 WUE 最低ꎬ表明其消耗等质量的
表皮有分布ꎬ这将有利于保持水分、减少蒸腾( 陈 水而积累的干物质最少ꎬ其对干旱环境的适应能
健辉等ꎬ2018)ꎻ没有分化出海绵组织和栅栏组织ꎬ 力可能相对较差ꎮ
这与叶庆生等(1992) 和李凤(2010) 对兰科植物 植物的光合参数反映不同植物对光环境的响
墨兰 ( Cymbidium sinense) 和 五 唇 兰 ( Phalaenopsis 应对策ꎬ可探讨植物对环境的适应能力( Yokoya et
pulcherrima)的叶片结构研究结果一致ꎮ 叶片厚度 al.ꎬ2007)ꎮ P max 越高ꎬ固碳能力越强ꎬ也有利于有
增大ꎬ能够促进叶片吸收更多的光能ꎬ提高光能利 机物的积累( Mahmud et al.ꎬ2018ꎻVincenza et al.ꎬ
 ̄2
用率(宋碧玉等ꎬ2017)ꎬ贵州地宝兰的叶片厚度和 2018)ꎮ 大花地宝兰的 P max (5.75 μmolm s )
 ̄1
叶肉厚度均显著(P<0.05)小于地宝兰和大花地宝 显著大于地宝兰和贵州地宝兰(分别为 4.09、3.89
兰ꎬ这可能是其光合能力较弱的原因之一ꎮ 较低 μmolm s )ꎬ说明大花地宝兰积累有机物的
 ̄1
 ̄2
的气孔密度及较大的气孔是植物在弱光环境下生 能力高于地宝兰和贵州地宝兰ꎮ LCP 和 LSP 反映
长的特性(盛洁悦等ꎬ2020)ꎮ 本研究中ꎬ大花地宝 植物光适应的生态幅度范围ꎬLCP 与 LSP 相差越
兰具有较低的气孔密度和较大的气孔面积ꎬ说明 大ꎬ植物对光的适应范围也越大(张旺锋等ꎬ2005)ꎮ