
Page 123 - 《广西植物》2020年第8期
P. 123
1 1 7 8 广 西 植 物 40 卷
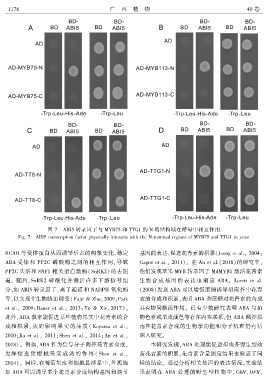
图 7 ABI5 转录因子与 MYB75 和 TTG1 的 N 端结构域在酵母中相互作用
Fig. 7 ABI5 transcription factor physically interacts with the N ̄terminal regions of MYB75 and TTG1 in yeast
RCAR 等受体蛋白从而诱导后者的构象变化ꎬ稳定 基因的表达ꎬ促进花青素的积累(Jeong et al.ꎬ 2004ꎻ
ABA 受体和 PP2C 磷酸酶之间的相互作用ꎬ导致 Gagné et al.ꎬ 2011)ꎮ 在 An et al.(2018)的研究中ꎬ
PP2C 失活和 SNF1 相关蛋白激酶(SnRK2) 的去阻 他们发现苹果 MYB 转录因子 MdMYB1 激活花青素
遏ꎮ 随后ꎬSnRK2 磷酸化并激活许多下游信号组 生物 合 成 基 因 的 表 达 也 响 应 ABAꎮ Loreti et al.
分ꎬ如 ABI5 转录因子、离子通道和 NADPH 氧化酶 (2008)发现 ABA 可以增强蔗糖诱导拟南芥中花青
等ꎬ以实现非生物胁迫耐受(Fujii & Zhuꎬ2009ꎻPark 素的合成和积累ꎬ表明 ABA 和蔗糖对花青素的合成
et al.ꎬ 2009ꎻBauer et al.ꎬ 2013ꎻYu & Xieꎬ2017)ꎮ 具有协同激活作用ꎮ 已有少数研究表明 ABA 与植
此外ꎬABA 激素能促进某些植物果实中花青素的合 物色素或果皮颜色等存在内在联系ꎬ但 ABA 调控拟
成和积累ꎬ 从而影响果实的品质 ( Koyama et al.ꎬ 南芥花青素合成的生物学功能和分子机理仍有待
2010ꎻJia et al.ꎬ 2011ꎻShen et al.ꎬ 2014ꎻAn et al.ꎬ 深入研究ꎮ
2018)ꎮ 例如ꎬABA 作为信号分子调控花青素合成ꎬ 本研究发现ꎬABA 处理能促进拟南芥野生型幼
发挥 促 进 甜 樱 桃 果 实 成 熟 的 作 用 ( Shen et al.ꎬ 苗花青素的积累ꎬ花青素含量测定结果也验证了同
2014)ꎮ 同样ꎬ在葡萄果皮和细胞悬浮系中ꎬ外源施 样的结论ꎮ 通过分析相关基因的表达情况ꎬ实验结
加 ABA 可以诱导多个花青素合成结构基因和调节 果表明在 ABA 处理的野生型植物中ꎬC4H、DFR、