
Page 101 - 《广西植物》2023年第10期
P. 101
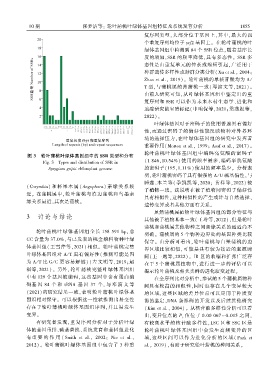
10 期 张梦洁等: 轮叶蒲桃叶绿体基因组特征及系统发育分析 1 8 5 5
复序列类型ꎬ大部分位于基因上ꎬ其中ꎬ最大的四
个重复序列均位于 ycf2 基因上ꎮ 在轮叶蒲桃的叶
绿体基因组中检测到 84 个 SSR 位点ꎬ随着基序长
度的增加ꎬSSR 的频率降低ꎬ具有多态性ꎮ SSR 多
态性是由重复单元的伸长或缩短引起ꎬ广泛用于
种群遗传多样性或种群分类分析(Xu et al.ꎬ 2004ꎻ
Zhao et al.ꎬ 2019)ꎮ 轮叶蒲桃的单核苷酸均为 A /
T 型ꎬ与蒲桃属的肖蒲桃一致( 邓演文等ꎬ2021)ꎮ
由前人研究可知ꎬ从叶绿体基因组中鉴定出的重
复序列和 SSR 可以作为未来木材生态学、进化和
追踪研究的宝贵标记(申响保等ꎬ2021ꎻ梁燕妮等ꎬ
2022)ꎮ
叶绿体基因对于密码子的使用普遍具有偏好
性ꎬ而通过密码子的偏好性能反映物种对外界环
境的选择压力ꎬ在叶绿体基因组的研究中发挥着
重要作用(Morton et al.ꎬ 1999ꎻ Asaf et al.ꎬ 2017)ꎮ
轮叶蒲桃叶绿体基因组中编码亮氨酸的密码子
图 3 轮叶蒲桃叶绿体基因组中的 SSR 类型和分布
(1 846ꎬ10.54%) 使用的频率最多ꎬ编码半胱氨酸
Fig. 3 Types and distribution of SSR in
Syzygium grijsii chloroplast genome 的密码子(195ꎬ1.11%) 使用的频率最少ꎮ 分析表
明ꎬ轮叶蒲桃密码子具有很强的 A / U 碱基偏性ꎬ与
睡莲、木兰等( 季凯凯等ꎬ2020ꎻ 黄祥等ꎬ2022) 被
(Corymbia) 和杯果木属 ( Angophora) 亲 缘 关 系 较
子植物一致ꎮ 这说明在被子植物间密码子偏好性
近ꎮ 在蒲桃属中ꎬ轮叶蒲桃与滇边蒲桃和乌墨亲
具有相似性ꎬ这种相似性的产生或许与自然选择、
缘关系最近ꎬ其次是蒲桃ꎮ
遗传变异或者其他方面有关系ꎮ
虽然蒲桃属植物叶绿体基因组的部分特征与
3 讨论与结论
其他被子植物基本一致(王玲等ꎬ2012)ꎬ但是轮叶
蒲桃和蒲桃属其他物种之间亲缘关系的远近尚不
轮叶蒲桃叶绿体基因组全长 158 591 bpꎬ总
明确ꎮ 蒲桃属的 5 个物种边界处的基因种类比较
GC 含量为 37.0%ꎬ与已发表的桃金娘科物种叶绿
保守ꎮ 由分析可看出ꎬ轮叶蒲桃与白果蒲桃的边
体基因组( 王雪芹等ꎬ2021) 相似ꎮ 轮叶蒲桃完整
界区域比较相似ꎬ可能是具有较为接近的起源时
叶绿体基因组对 A / T 具有偏好性( 推测可能是因
间(王一麾等ꎬ2022)ꎮ IR 区的收缩和扩张广泛存
为 A / T 比 G / C 更容易解链) ( 左文明等ꎬ2019ꎻ赵 在于 5 个蒲桃属植物中ꎬ进行进一步的评估可以
祺等ꎬ2021)ꎮ 另外ꎬ轮叶蒲桃完整叶绿体基因组
揭示轮叶蒲桃及相关类群的进化演变过程ꎮ
中有 129 个基因被编码ꎬ这些基因中含有蛋白编 在全序列比对分析中ꎬ参试的 5 个蒲桃属物种
码基 因 84 个 和 tRNA 基 因 37 个ꎬ 与 邓 演 文 等 间具有较高的相似性ꎬ同时也存在几个变异较大
(2021)的研究结果一致ꎬ表明轮叶蒲桃叶绿体基 的区域ꎬ这些区域的差异位点可以应用于种质资
因组相对保守ꎮ 可以根据这一性状推测出易变性 源的鉴定、DNA 条形码的开发以及后续其他研究
存在于轮叶蒲桃叶绿体基因组序列ꎬ并且易发生 (Kim et al.ꎬ 2004)ꎮ 从核苷酸多样性分析可以看
变异ꎮ 出ꎬ变异位点的 P 值位于 0.00 067 ~ 0.015 之间ꎬ
i
有研究者发现ꎬ重复序列分析对于分析叶绿 有较高水平的核苷酸多样性ꎬLSC 区和 SSC 区是
体的基因重排、碱基替换、系统发育和基因组进化 轮叶蒲桃叶绿体基因组中会发生高频变异的区
有重 要 的 作 用 ( Smith et al.ꎬ 2002ꎻ Nie et al.ꎬ 域ꎬ这些区间可以作为进化分析的区域( Park et
2012)ꎮ 轮叶蒲桃叶绿体基因组中包含了 3 种重 al.ꎬ 2019)ꎬ有助于研究轮叶蒲桃的种间关系ꎮ