
Page 162 - 《广西植物》2023年第10期
P. 162
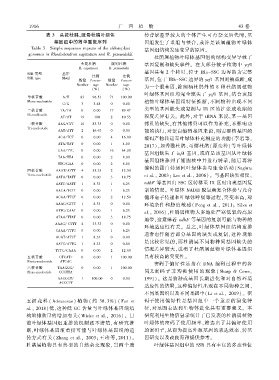
1 9 1 6 广 西 植 物 43 卷
表 3 头花杜鹃、陇蜀杜鹃叶绿体 传背景差异较大的个体产生可育杂交后代时ꎬ基
基因组中的简单重复序列 因组发生了重组与整合ꎬ或许是该属植物叶绿体
Table 3 Simple sequence repeats of the chloroplast
基因组结构发生变异的原因ꎮ
genomes in Rhododendron capitatum and R. przewalskii
杜鹃属植物叶绿体基因组的结构变异导致了
头花杜鹃 陇蜀杜鹃 基因复制和缺失事件ꎮ 在大部分被子植物中 ycf1
R. capitatum R. przewalskii
基因具有 2 个拷贝ꎬ位于 IRa-SSC 边界的为完整
SSR 类型 基序
比例 比例
SSR type Motif 基因ꎬ位于 IRb-SSC 边界的 ycf1 基因则被截断ꎬ成
数量 Percen ̄ 数量 Percen ̄
Number tage Number tage 为一个假基因ꎬ除阔柄杜鹃外的 8 种杜鹃属植物
(%) (%)
叶绿体基因组均完全缺失了 ycf1 基因ꎬ结合该属
单核苷酸 A / T 83 96.51 71 100.00
Mono ̄nucleotide 植物叶绿体基因组特征推断ꎬ不同物种出现不同
C / G 3 3.48 0 0.00
类型的基因缺失或复制与 IR 区的扩张或收缩的
二核苷酸 TA / TA 0 0.00 17 89.47
Di ̄nucleotide 程度差异有关ꎮ 此外ꎬ对于 tRNA 来说ꎬ某一基因
AT / AT 19 100 2 10.53
三核苷酸 AAG / CTT 10 83.33 0 0.00 拷贝的缺失ꎬ有其他拷贝可以作为补充ꎬ不影响功
Tri ̄nucleotide
AAT / ATT 2 16.67 0 0.00 能的执行ꎬ对蛋白编码基因来说ꎬ则需要核基因的
AGA / TCT 0 0.00 4 16.00
编码产物转运至叶绿体补充相应的功能( 李岩等ꎬ
2017)ꎬ如秀雅杜鹃、马缨杜鹃( 原变种) 等叶绿体
ATA / TAT 0 0.00 1 4.00
GAA / TTC 0 0.00 16 64.00
基因组缺失了 infA 基因ꎬ或许是该基因从叶绿体
TAA / TTA 0 0.00 2 8.00
基因组转移到了细胞核中并进行转录ꎬ随后再将
TTC / GAA 0 0.00 2 8.00
编码的蛋白转运回叶绿体参与生命活动( Sugiura
四核苷酸 AAAT / ATTT 4 33.33 2 12.50
Tetra ̄nucleotide et al.ꎬ 2003ꎻ Lee et al.ꎬ 2006)ꎮ 与基因缺失相反ꎬ
AATA / TATT 0 0.00 3 18.75
ndhF 等基因由 SSC 区转移至 IR 区时出现基因复
AATT / AATT 1 8.33 1 6.25
AAGA / TCTT 0 0.00 1 6.25 制的情况ꎮ 叶绿体 NADH 脱氢酶复合体参与光合
循环电子传递和叶绿体呼吸等过程ꎬ突变率高ꎬ对
AGAA / TTCT 0 0.00 2 12.50
AAAC / GTTT 1 8.33 0 0.00 环境条件和胁迫敏感( Peng et al.ꎬ 2011ꎻ Silva et
ATTG / CAAT 0 0.00 1 6.25 al.ꎬ 2016)ꎬ杜鹃属植物大多地处严寒低氧的高原
ATAA / TTAT 0 0.00 3 18.75
地带ꎬ这意味着 ndhF 等基因的复制可能与物种的
AAAG / CTTT 4 33.33 0 0.00
环境适应性有关ꎮ 总之ꎬ叶绿体基因组结构重排
CAAA / TTTG 0 0.00 1 6.25
通常也伴随着部分基因的缺失或复制ꎬ这种现象
AGAT / ATCT 1 8.33 0 0.00
是比较常见的ꎬ而杜鹃属不同物种间基因缺失的
AATC / ATTG 1 8.33 0 0.00
情况差异较大ꎬ说明了杜鹃属植物叶绿体基因组
TTTC / GAAA 0 0.00 2 12.50
五核苷酸 GTCAT / 0 0.00 1 100.00 具有较高的突变性ꎮ
Penta ̄nucleotide ATGAC
密码子偏好性是指在 DNA 编码过程中的各
六核苷酸 TAAGGG / 0 0.00 1 100.00
Hexa ̄nucleotide CCCTTA 同义密码 子 非 均 衡 使 用 的 现 象 ( Sharp & Coweꎬ
1991)ꎮ 这是物种或基因长期进化和对自然环境
AAGGGT / 1 100.00 0 0.00
ACCCTT
适应性的结果ꎬ这种偏好性出现在不同物种之间、
不同基因组以及不同基因中( Li et al.ꎬ 2019)ꎮ 密
五福 花 科 ( Adoxaceae) 植 物 ( 约 38. 3%) ( Fan et 码子使用偏好性是基因组中一个重要的演化特
al.ꎬ 2018)低ꎬ这种低 GC 含量与叶绿体基因组结 征ꎬ对基因表达和生物体进化具有重要意义ꎮ 本
构重排数量的增加有关( Wicke et al.ꎬ 2016)ꎮ 目 研究利用生物信息学统计了已发表的杜鹃属植物
前叶绿体基因组重排的机制还不清楚ꎬ有研究推 叶绿体的密码子使用频率ꎬ筛选出了其偏好使用
断ꎬ叶绿体基因组重排可能与叶绿体基因组的遗 的密码子ꎬ从而为提高外源基因的表达效率、转基
传方式有关( Zhang et al.ꎬ 2003ꎻ王玲等ꎬ2011)ꎮ 因研究以及改良育种提供参考ꎮ
杜鹃属植物具有典型的自然杂交现象ꎬ当两个遗 叶绿体基因组中的 SSR 具有丰富的多态性位