
Page 148 - 广西植物2024年1期
P. 148
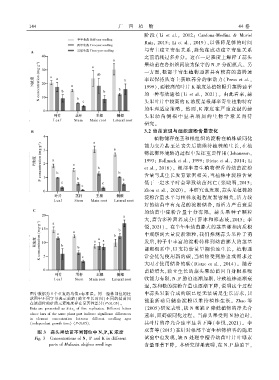
1 4 4 广 西 植 物 44 卷
阶 段 ( Li et al.ꎬ 2012ꎻ Cardona ̄Medina & Muriel
Ruizꎬ 2015ꎻ Li et al.ꎬ 2019)ꎬ以保障足够的时间
与寄主建立寄生关系ꎬ避免在成功建立寄生关系
之前消耗过多养分ꎮ 这在一定程度上解释了蒜头
果幼苗在各组织间较为保守的 N、P 分配模式ꎮ 另
一方面ꎬ根部半寄生植物通常具有较高的蒸腾速
率以保持从寄主获取养分的驱动力( Press et al.ꎬ
1999)ꎬ而较高的叶片 K 浓度是植物提升蒸腾速率
的一种有效途径( Li et al.ꎬ 2021)ꎮ 由此看来ꎬ蒜
头果叶片中较高的 K 浓度是根部半寄生植物特有
的生理适应策略ꎮ 然而ꎬK 浓度在严重衰退的蒜
头果 幼 苗 侧 根 中 显 著 增 加 的 生 物 学 意 义 尚 待
研究ꎮ
3.2 幼苗衰退与组织淀粉含量变化
植物储存在茎和根组织的淀粉在植株碳同化
能力变差甚至是丧失后能维持植株的生长ꎬ在植
物抵御环境胁迫过程中发挥重要作用( Johanssonꎬ
1993ꎻ Bollmark et al.ꎬ 1999ꎻ Dietze et al.ꎬ 2014ꎻ Li
et al.ꎬ 2018)ꎮ 根部半寄生植物檀香的幼苗淀粉
含量与其生长发育紧密相关ꎬ当植株中淀粉含量
低于一定水平时会导致幼苗死亡( 张晓明ꎬ2015ꎻ
Zhou et al.ꎬ 2020)ꎮ 本研究也发现ꎬ蒜头果植株的
淀粉含量水平与植株衰退程度紧密相关ꎬ活力较
好的幼苗中有充足的淀粉储备ꎬ而活力严重衰退
的幼苗中淀粉含量十分有限ꎮ 蒜头果种子颗粒
大ꎬ富含多种营养成分( 薛冰和邵志凌ꎬ2015ꎻ 李
悦ꎬ2021)ꎮ 在半年生幼苗膨大的茎基部和肉质根
中观察到大量淀粉颗粒ꎬ我们推测蒜头果种子萌
发后ꎬ种子中丰富的淀粉转移到幼苗膨大的茎基
部和根系中ꎬ以支持幼苗早期快速生长ꎮ 植物通
常会优先使用新的碳ꎬ当植物受到胁迫或需求过
大时才使用储备的碳( Dietze et al.ꎬ 2014)ꎮ 随着
苗龄增大ꎬ独立生长的蒜头果幼苗因自身根系吸
收能力有限ꎬN、P 胁迫逐渐加剧ꎬ导致植株逐渐衰
退ꎬ茎和根的淀粉含量也逐渐下降ꎬ说明这个过程
图中数据为 5 个重复的均值±标准误ꎮ 同一植株部位的柱 中蒜头果新合成的碳已经无法满足生长需求ꎬ只
状图中不同字母表示苗龄(独立生长时间) 不同的幼苗间 能重新动员储备淀粉以维持植株生长ꎮ Zhao 等
在该部位的矿质元素浓度存在显著性差异(P<0.05)ꎮ
Data are presented as x ± s x of five replicates. Different letters (2005)研究表明ꎬ缺 N 和缺 P 降低植物的净光合
above bars of the same plant part indicate significant differences
速率ꎬ阻碍碳同化过程ꎮ 当蒜头果受到 N 胁迫时ꎬ
in element concentration between different seedling ages
(independent growth time) (P<0.05). 其叶片的净光合速率显著下降( 李悦ꎬ2021)ꎮ 李
双喜等(2015)在针对根部半寄生植物檀香的施肥
图 3 蒜头果幼苗不同部位中 N、P、K 浓度
试验中也发现ꎬ缺 N 处理中檀香幼苗叶片叶绿素
Fig. 3 Concentrations of Nꎬ P and K in different
parts of Malania oleifera seedlings 含量显著下降ꎮ 本研究结果表明ꎬ在 N、P 胁迫下ꎬ