
Page 205 - 《广西植物》2024年第5期
P. 205
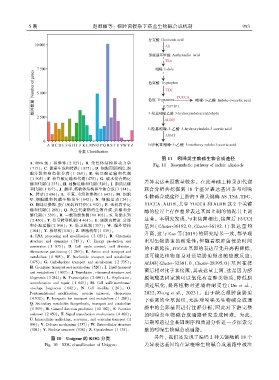
5 期 赵淑丽等: 根肿菌侵染下菘蓝生物碱合成机制 9 9 3
图 11 吲哚类生物碱生物合成途径
A. RNA 加 工 和 修 饰 ( 2 023)ꎻ B. 染 色 体 结 构 和 动 力 学 Fig. 11 Biosynthetic pathway of indole alkaloids
(715)ꎻ C. 能量生成和转换(1 875)ꎻ D. 细胞周期调控、细
胞分 裂 和 染 色 体 分 离 ( 1 269)ꎻ E. 氨 基 酸 运 输 和 代 谢
(1 905)ꎻ F. 核苷酸运输和代谢(475)ꎻ G. 碳水化合物运
输和代谢(2 335)ꎻ H. 辅酶运输和代谢(520)ꎻ I. 脂质运输 差异表达基因数量较多ꎮ 在此基础上转录加代谢
和代谢(1 897)ꎻ J. 翻译、核糖体结构和生物合成(3 214)ꎻ 联合分析共挖掘到 18 个差异表达基因参与吲哚
K. 转录(2 686)ꎻ L. 重复、重组和修饰(1 643)ꎻ M. 细胞
生物碱合成途径上游的 5 种关键酶 AS、TSA、TDC、
壁、细胞膜 和 包 膜 生 物 发 生 ( 642)ꎻ N. 细 胞 运 动 ( 24)ꎻ
O. 翻译后修饰、蛋白反转和伴侣(4 932)ꎻ P. 无机离子运 YUCCA、ALDHꎬ其中 YUCCA 和 ALDH 这 2 个关键
输和代谢(1 288)ꎻ Q. 次生代谢物的生物合成、运输和分 酶的位置上存在差异表达基因上调的情况且上调
解代谢(1 559)ꎻ R. 一般功能预测(10 102)ꎻ S. 功能未知
(2 450)ꎻ T. 信号转导机制(4 461)ꎻ U. 细胞内转运、分泌 显著ꎮ 本研究发现ꎬ与未接菌相比ꎬ接菌后 YUCCA
和小泡运输(2 096)ꎻ V. 防卫机制(357)ꎻ W. 胞外结构 基因 ( Cluster ̄36192. 0、 Cluster ̄36192. 1) 表 达 量 均
(104)ꎻ Y. 核结构(138)ꎻ Z. 细胞构架(1 135)ꎮ
升高ꎬ这与 Cao 等(2019)的研究结果一致ꎬ都呈现
A. RNA processing and modification ( 2 023)ꎻ B. Chromatin
出对生物胁迫的耐受性ꎻ但随着根肿菌侵染时间
structure and dynamics ( 715 )ꎻ C. Energy production and
conversion ( 1 875 )ꎻ D. Cell cycle controlꎬ cell divisionꎬ
的不断延长ꎬYUCCA 基因的表达量先升高再降低ꎬ
chromosome partitioning (1 269)ꎻ E. Amino acid transport and
metabolism ( 1 905)ꎻ F. Nucleotide transport and metabolism 这可能是植物自身对逆境胁迫做出的应激反应ꎻ
ALDH( Cluster ̄32381.0、Cluster ̄28395.0) 基因在接
(475)ꎻ G. Carbohydrate transport and metabolism ( 2 335)ꎻ
H. Coenzyme transport and metabolism (520)ꎻ I. Lipid transport
and metabolism (1 897)ꎻ J. Translationꎬ ribosomal structure and 菌后相对比于未接菌ꎬ其表达量上调ꎬ这是因为醛
biogenesis (3 214)ꎻ K. Transcription (2 686)ꎻ L. Replicationꎬ 脱氢酶基因家族可以氧化有毒醛类物质ꎬ降低脂
recombination and repair ( 1 643)ꎻ M. Cell wall/ membrane/
质过氧化ꎬ提高植物对逆境的耐受性( Du et al.ꎬ
envelope biogenesis ( 642 )ꎻ N. Cell motility ( 24 )ꎻ O.
Posttranslational modificationꎬ protein turnoverꎬ chaperones 2022ꎻZhang et al.ꎬ 2023)ꎮ 由于缺乏根肿菌胁迫
(4 932)ꎻ P. Inorganic ion transport and metabolism (1 288)ꎻ 下菘蓝的全基因组ꎬ无法对吲哚类生物碱合成通
Q. Secondary metabolite biosynthesisꎬ transport and catabolism
路中的全部基因进行注释分析ꎬ因此对下游完整
(1 559)ꎻ R. General function prediction (10 102)ꎻ S. Function
unknown (2 450)ꎻ T. Signal transduction mechanisms (4 461)ꎻ 的吲哚类生物碱合成通路研究造成困难ꎮ 为此ꎬ
U. Intracellular traffickingꎬ secretionꎬ and vesicular transport (2
后期可通过全基因测序和质谱分析进一步探索完
096)ꎻ V. Defense mechanism (357)ꎻ W. Extracellular structure
(104)ꎻ Y. Nuclear structure (138)ꎻ Z. Cytoskeleton (1 135). 整的吲哚生物碱合成通路ꎮ
图 10 Unigene 的 KOG 分类 另外ꎬ 我们还发现了编码 2 种关键酶的 18 个
Fig. 10 KOG classification of Unigene 差异表达基因均在异喹啉生物碱合成通路中被注