
Page 130 - 《广西植物》2024年第7期
P. 130
1 3 3 0 广 西 植 物 44 卷
∗表示差异显著(P<0.05)ꎮ
∗ indicates significant differences (P<0.05).
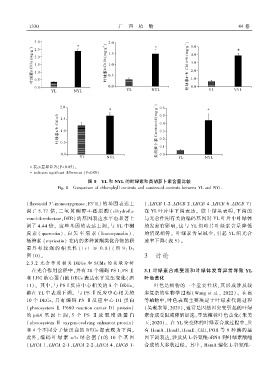
图 8 YL 和 NYL 的叶绿素和类胡萝卜素含量比较
Fig. 8 Comparison of chlorophyll contents and carotenoid contents between YL and NYL
(flavonoid 3′ ̄monooxygenaseꎬF3′H) 的基因表达上 1、LHCB 1 ̄2、LHCB 2、LHCB 4、LHCB 6、LHCB 7)
调了 5. 77 倍ꎬ 二 氢 黄 酮 醇 ̄4 ̄还 原 酶 ( dihydrofla ̄ 在 YL 叶片中下调表达ꎮ 综上结果表明ꎬ下调的
vonol ̄4 ̄reductaseꎬDFR) 的基因表达水平也显著上 与光合作用有关的编码基因对 YL 叶片中叶绿体
调了 4.44 倍ꎮ 这些基因的表达上调ꎬ与 YL 中槲 的发育有影响ꎬ这与 YL 组叶片叶绿素含量降低
皮 素 ( quercetin)、 白 矢 车 菊 素 ( leucocyanidin )、 的情况相符ꎮ 叶绿素含量减少ꎬ引起 YL 组光合
杨梅素 (myricetin) 在内的多种黄酮类化合物的积 速率下降( 表 5) ꎮ
累具 有 较 强 的 相 关 性 ( | r | ≥ 0. 8) ( 图 9: Dꎻ
图 10)ꎮ 3 讨论
2.3.2 光合作用相关 DEGs 和 SCMs 的关联分析
在光合作用途径中ꎬ共有 28 个编码 PS I、PS Ⅱ 3.1 叶绿素合成受阻和叶绿体发育异常导致 YL
和 LHC 核心蛋白的 DEGs 表达水平发生变化( 图 叶色黄化
11) ꎮ 其中ꎬ与 PS I 反应中心相关的 8 个 DEGsꎬ 叶色是植物的一个重要性状ꎬ其形成涉及较
都在 YL 中表现下调ꎮ 与 PS Ⅱ反应中心相关的 多复杂的生物学过程( Wang et al.ꎬ 2022)ꎮ 在高
10 个 DEGsꎬ 只 有 编 码 PS Ⅱ 反 应 中 心 D1 蛋 白 等植物中ꎬ叶色表现主要决定于叶绿素代谢过程
( photosystem Ⅱ P680 reaction center D1 protein) (吴砚农等ꎬ2021)ꎬ通常是因基因突变引起的叶绿
的 psbA 基 因 上 调ꎬ 5 个 PS Ⅱ 放 氧 增 强 蛋 白 素合成受阻或降解加速ꎬ导致植株叶色黄化( 朱美
( photosystem Ⅱ oxygen ̄evolving enhancer protein) 玉ꎬ2020)ꎮ 在 YL 突变体的叶绿素合成过程中ꎬ共
和 4 个不同分子量蛋白的 DEGs 都表现为下调ꎮ 有 HemA、HemB、HemE、ChlI、POR 等 9 种酶的基
此外ꎬ 编 码 叶 绿 素 a / b 结 合 蛋 白 的 10 个 基 因 因下调表达ꎬ涉及从 L ̄谷氨酰 ̄tRNA 到叶绿素酸酯
( LHCA 1、LHCA 2 ̄1、LHCA 2 ̄2、LHCA 4、LHCB 1 ̄ 合成的大多数过程ꎮ 其中ꎬ HemA 催化 L ̄谷氨酰 ̄