
Page 130 - 《广西植物》2020年第1期
P. 130
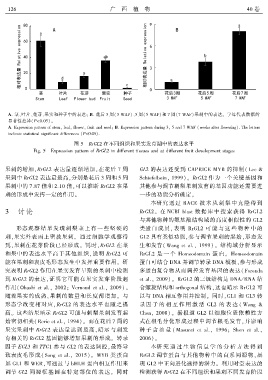
1 2 6 广 西 植 物 40 卷
A. 茎、叶片、花芽、果实和种子中的表达ꎻ B. 花后 3 周(3 WAF)、5 周(5 WAF)和 7 周(7 WAF)果刺中的表达ꎮ 字母代表数据的
显著性差异(P<0.05)ꎮ
A. Expression pattern of stemꎬ leafꎬ flowerꎬ fruit and seedꎻ B. Expression pattern during 3ꎬ 5 and 7 WAF (weeks after flowering). The letters
indicate statistical significant differences (P<0.05).
图 5 RrGL2 在不同组织和果实发育期中的表达水平
Fig. 5 Expression pattern of RrGL2 in different tissues and at different fruit development stages
果刺的增加ꎬRrGL2 表达量逐渐增加ꎬ在花后 7 周 GL2 的表达还受到 CAPRICE MYB 的抑制( Lee &
果刺中 RrGL2 表达量最高ꎬ分别是花后 3 周和 5 周 Schiefelbeinꎬ 1999)ꎮ RrGL2 作为一个关键基因和
果刺中的 7.87 倍和 2.10 倍ꎬ可以推断 RrGL2 在果 其他参与调节刺梨果刺发育的基因功能还需要进
刺的形成中发挥一定的作用ꎮ 一步的功能分析确定ꎮ
本研究 通 过 RACE 技 术 从 刺 梨 中 克 隆 得 到
3 讨论 RrGL2ꎮ 在 NCBI blast 数 据 库 中 搜 索 获 得 RrGL2
与其他物种的氨基酸结构域的高度相似性的 GL2
形态观察结果发现刺 梨 茎 上 有 一 些 坚 硬 的 类蛋白 成 员ꎬ 表 明 RrGL2 可 能 与 这 些 物 种 中 的
刺ꎬ果实外表面上密披果刺ꎮ 通过细胞学观察得 GL2 具有类似功能ꎬ参与调节果刺的起始ꎬ形态发
到ꎬ果刺在花芽阶段已经形成ꎮ 同时ꎬRrGL2 在茎 生和发育( Wang et al.ꎬ 1999)ꎮ 结构域分析显示
和果中的表达水平高于其他组织ꎬ说明 RrGL2 可 RrGL2 是 一 个 Homeodomain 蛋 白ꎮ Homeodomain
能在果刺和表皮毛形态发生中发挥重要作用ꎮ 研 蛋白可结合 DNA 并调节转录 DNA 模板ꎬ参与形成
究表明 RrGL2 作用在果实发育早期的果刺中检测 多蛋白复合物从而调控发育基因的表达( Foronda
到 RrGL2 的表达ꎬ证明它可能在果实发芽阶段起 et al.ꎬ 2009)ꎮ RrGL2 的三级结构是 DNA / RNA 结
作用(Ohashi et al.ꎬ 2002ꎻ Vernoud et al.ꎬ 2009)ꎮ 合螺旋结构和 orthogonal 结构ꎬ这也暗示 RrGL2 可
随着果实的成熟ꎬ果刺的数量和长度都增加ꎮ 与 以与 DNA 相互作用并控制ꎮ 同时ꎬGL1 和 GL3 转
形态学改变相对应ꎬRrGL2 的表达水平也随之提 录因 子 的 相 互 作 用 激 活 GL2 的 表 达 ( Wang &
高ꎮ 这些结果暗示 RrGL2 可能与刺梨果刺发育起 Chenꎬ 2008)ꎮ 据报道 GL2 以细胞位置依赖性方
始密切相关(Rerie et al.ꎬ 1994)ꎮ 而在花后 7 周的 式在根毛分化形成过程中调节根毛发育ꎬ并影响
果实果刺中 RrGL2 表达量达到最高ꎬ暗示与刺发 种 子 含 油 量 ( Masucci et al.ꎬ 1996ꎻ Shen et al.ꎬ
育相关的 RrGL2 基因能够增加果刺的形成ꎮ 转录 2006)ꎮ
因子 EGL3 和 TTG1 参与 GL2 的表达调控ꎬ最终导 本研 究 通 过 生 物 信 息 学 的 分 析 方 法 得 到
致表皮毛形成( Song et al.ꎬ 2015)ꎮ MYB 类蛋白 RrGL2 调节蛋白与其他物种中的直系同源物ꎬ展
如 GL1 和 WERꎬ可通过与 bHLH 蛋白相互作用来 现 GL2 中正向进化选择的潜力ꎮ 利用时空表达的
调节 GL2 同源框基因在特定部位的表达ꎮ 同时 检测获得 RrGL2 在不同组织和果刺不同发育阶段