
Page 172 - 《广西植物》2023年第10期
P. 172
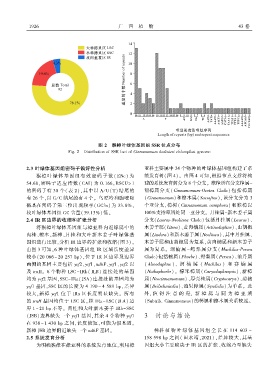
1 9 2 6 广 西 植 物 43 卷
图 2 猴樟叶绿体基因组 SSR 位点分布
Fig. 2 Distribution of SSR loci of Cinnamomum bodinieri chloroplast genome
2.3 叶绿体基因组密码子偏好性分析 亚科主要属中 24 个物种的叶绿体基因组构建了系
猴樟 叶 绿 体 基 因 组 有 效 密 码 子 数 ( ENc) 为 统发育树(图 4)ꎮ 由图 4 可知ꎬ根据节点支撑将构
54.68ꎬ密码子适应指数( CAI) 为 0.166ꎬRSCU > 1 建的系统发育树分为 8 个分支ꎮ 猴樟所在分支樟属-
的密码子有 30 个( 表 2)ꎬ其中以 A / U( T) 结尾的 甜樟属分支( Cinnamomum ̄Ocotea Clade) 包括樟属
有 26 个ꎬ以 G / C 结尾的有 4 个ꎮ 鸟嘌呤和胞嘧啶 (Cinnamomum)和檫木属(Sassafras)ꎬ该分支分为 3
碱基在密码子第三位出现频率( GC3s) 为 33.8%ꎬ 个亚分支ꎬ樟树( Cinnamomum camphora) 和猴樟以
较叶绿体基因组 GC 含量(39.13%)低ꎮ 100%支持率所处同一亚分支ꎮ 月桂属-新木姜子属
2.4 IR 区边界的收缩和扩张分析 分支(Laurus ̄Neolitsea Clade) 包括月桂属(Laurus)、
将猴樟叶绿体基因组与樟亚科内近缘属中的 木姜子属(Litsea)、黄肉楠属(Actinodaphne)、山胡椒
肉桂、檫木、新樟、月桂和大叶新木姜子叶绿体基 属(Lindera)和新木姜子属(Neolitsea)ꎬ其中月桂属、
因组进行比较ꎬ分析 IR 边界的扩张和收缩(图 3)ꎮ 木姜子属和山胡椒属为复系ꎬ黄肉楠属和新木姜子
由图 3 可知ꎬ6 种叶绿体基因组 IR 区域长度差异 属为复系ꎮ 润楠属 - 鳄梨属分支 ( Machilus ̄Persea
较小(20 066 ~ 20 257 bp)ꎬ位于 IR 区边界及边界 Clade)包括楠属(Phoebe)、鳄梨属(Persea)、油丹属
两侧的基因主要包括 ycf2、ycf1、ndhF、ycf1、ycf2 以 ( Alseodaphne )、 润 楠 属 ( Machilus ) 和 赛 楠 属
及 trnHꎮ 6 个物种 LSC -IRb( JLB) 连接处的基因 (Nothaphoebe)ꎮ 檬果樟属( Caryodaphnopsis)、新樟
均为 ycf2 基因ꎬSSC-IRa( JSA) 连接处的基因均为 属(Neocinnamomum)、厚壳桂属(Cryptocarya)、琼楠
ycf1 基因ꎬSSC 区的长度为 4 190 ~ 4 589 bpꎬ差异 属(Beilschmiedia)、油果樟属(Syndiclis) 为单系ꎮ 此
较大ꎬ新樟 ycf1 位于 IRa 区长度明显缺失ꎮ 所有 外ꎬ 值 得 注 意 的 是ꎬ 新 樟 属 与 同 为 樟 亚 族
的 trnH 基因均位于 LSC 区ꎬ距 IRa-LSC( JLA) 边 (Subtrib. Cinnamomeae)的樟属和檫木属关系较远ꎮ
界 1 ~ 21 bp 不等ꎮ 肉桂和大叶新木姜子 IRb-SSC
(JSB)边界缺失一个 ycf1 基因ꎬ其余 4 个物种 ycf1 3 讨论与结论
在 938 ~ 1 430 bp 之间ꎬ长度较短ꎬ可能为假基因ꎮ
新樟 JSB 边界附近缺失一个 ndhF 基因ꎮ 樟科 植 物 叶 绿 体 基 因 组 全 长 在 114 603 ~
2.5 系统发育分析 158 598 bp 之间( 田永靖ꎬ2021)ꎬ差异较大ꎬ其基
为明确猴樟在樟亚科的系统发育地位ꎬ利用樟 因组大小主要取决于 IR 区的扩张、收缩乃至缺失