
Page 126 - 《广西植物》2023年第7期
P. 126
1 2 9 4 广 西 植 物 43 卷
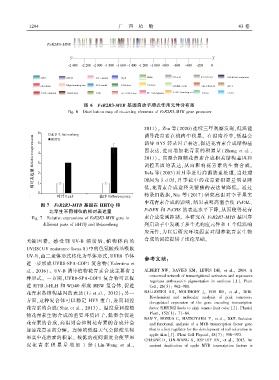
图 6 FeR2R3 ̄MYB 基因启动子顺式作用元件分布图
Fig. 6 Distribution map of cis ̄acting elements of FeR2R3 ̄MYB gene promoter
2011)ꎮ Zhu 等(2020)连续三年观察发现ꎬ低温能
诱导花青素在桃肉中积累ꎮ 在拟南芥中ꎬ低温会
诱导 HY5 转录因子表达ꎬ促进花青素合成结构基
因表达ꎬ进而增加花青素的积累量( Zhang et al.ꎬ
2011)ꎮ 高温会抑制花青素合成相关结构基因和
调控基因的表达ꎬ从而影响花青素的生物合成ꎮ
Dela 等(2003)对月季进行高温胁迫处理ꎬ当处理
时间为 3 d 时ꎬ月季花中的花青素积累量明显降
低ꎬ花青素合成途径关键酶的表达量降低ꎮ 通过
转录组数据ꎬNiu 等(2017) 研究高温对李子果实
中花青素合成的影响ꎬ结果表明高温会使 PsPAL、
图 7 FeR2R3 ̄MYB 基因在 HHTQ 和
北早生不同部位的相对表达量 PsDFR 和 PsCHS 的表达水平下降ꎬ从而使得花青
素合成受到抑制ꎮ 本研究在 FeR2R3 ̄MYB 基因序
Fig. 7 Relative expressions of FeR2R3 ̄MYB gene in
different parts of HHTQ and Beizaosheng 列启动子中发现了多个光响应元件和 1 个低温响
应元件ꎬ为以后研究环境因素对甜荞花青素生物
关键 因 素ꎮ 感 受 到 UV ̄B 辐 射 后ꎬ 植 物 体 内 的 合成的调控提供了理论基础ꎮ
UVR8(UV resistance locus 8)中的色氨酸残基吸收
UV ̄Bꎬ由二聚体形式转化为单体形式ꎬUVR8 单体 参考文献:
进一步形成 UVR8 ̄SPA ̄COP1 复合物( Valentina et
al.ꎬ 2016)ꎮ UV ̄B 诱导植物花青素合成主要有 2 ALBERT NWꎬ DAVIES KMꎬ LEWIS DHꎬ et al.ꎬ 2014. A
种形式ꎮ 一方面ꎬUVR8 ̄SPA ̄COP1 复合物可以促 conserved network of transcriptional activators and repressors
regulates anthocyanin pigmentation in eudicots [ J]. Plant
进 MYB、bHLH 和 WD40 形成 MBW 复合体ꎬ促进 Cellꎬ 26(3): 962-980.
花青素各结构基因的表达( Li et al.ꎬ 2012)ꎻ另一 BALLESTER ARꎬ MOLTHOFF Jꎬ VOS RDꎬ et al.ꎬ 2010.
Biochemical and molecular analysis of pink tomatoes:
方面ꎬ这种复合体可以稳定 HY5 蛋白ꎬ进而调控
deregulated expression of the gene encoding transcription
花青素的合成(Shin et al.ꎬ 2013)ꎮ 温度是调控植 factor SlMYB12 leads to pink tomato fruit color [J]. Physiol
物花青素生物合成的重要环境因子ꎬ低温会促进 Plantꎬ 152(1): 71-84.
BAN Yꎬ HONDA Cꎬ HATSUYAMA Yꎬ et al.ꎬ 2007. Isolation
花青素的合成ꎬ高温则会抑制花青素的合成并会 and functional analysis of a MYB transcription factor gene
加速花青素的分解ꎮ 连续的低温天气会促使果树 that is a key regulator for the development of red coloration in
apple skin [J]. Plant Cell Physiolꎬ 48(7): 958-970.
果实中花青素的积累ꎮ 较低的夜间温度会使苹果
CHAGNÉ Dꎬ LIN ̄WANG Kꎬ ESPLEY RVꎬ et al.ꎬ 2013. An
皮花 青 素 积 累 量 增 加 1 倍 ( Lin ̄Wang et al.ꎬ ancient duplication of apple MYB transcription factors is