
Page 115 - 《广西植物》2020年第2期
P. 115
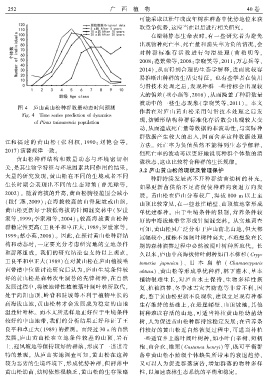
2 5 2 广 西 植 物 40 卷
可能采取以壮年或成年树在群落中优势地位来获
取竞争优势ꎬ这应当在以后进行相关研究ꎮ
在编制静态生命表时ꎬ有一些研究者为避免
出现物种死亡率、死亡量和消失率为负的情况ꎬ会
对种 群 标 准 存 活 数 进 行 匀 滑 处 理 ( 苗 艳 明 等ꎬ
2008ꎻ范繁荣等ꎬ2008ꎻ李晓笑等ꎬ2011ꎻ万志兵等ꎬ
2014)ꎬ从而得到合理的生态学解释ꎬ进而比较容
易推断出种群的生活史特征ꎮ 也有些学者在使用
匀滑技术处理之后ꎬ发现种群一些指标会出现较
大的偏差(项小燕等ꎬ2016)ꎬ从而掩盖了种群数量
波动中的一些生态现象( 李晓笑等ꎬ2011)ꎮ 本文
图 4 庐山黄山松种群数量动态时间预测
作者在对庐山黄山松采用匀滑技术处理之后发
Fig. 4 Time series prediction of dynamics
现ꎬ纺锤形结构种群标准化存活数会出现较大变
of Pinus taiwanensis population
动ꎬ从而造成死亡量等数据的非波动性ꎬ与实际种
群数据产生较大的出入ꎬ因而舍弃这种数据处理
江和 福 建 的 黄 山 松 ( 张 利 权ꎬ 1990ꎻ 刘 艳 会 等ꎬ
方法ꎮ 死亡率为负值虽然不能得到生态学解释ꎬ
2017)演替规律一致ꎮ
但死亡率的波动可以更好地说明种群个体数的消
黄山松种群结构和数 量 动 态 与 环 境 密 切 相
涨状态ꎬ这也比较符合种群的生长规律ꎮ
关ꎬ是其生物学特征与环境因素共同作用的结果ꎮ
3.2 庐山黄山松的现状及管理保护
大量的研究发现ꎬ黄山松在不同的生境或者不同
种群持续发展离不开种群幼苗幼树的补充ꎬ
生长时期会表现出不同的生态对策( 曹光球等ꎬ
如果更新苗供给不足将促使种群向衰退方向发
2002)ꎮ 随着海拔的升高ꎬ黄山松胸径范围会减小 展ꎮ 黄山松在庐山分布较广ꎬ海拔 800 m 以上至
(段仁燕ꎬ2009)ꎻ在海拔较高的山脊陡坡或山顶ꎬ 山顶比较常见ꎬ在一些悬崖峭壁、山顶坡地常形成
黄山松更新好于较低海拔的针阔混交林中( 罗世 单优建群种ꎮ 由于生境条件的限制ꎬ营养条件较
家等ꎬ1999ꎻ李家湘等ꎬ2004)ꎻ较高海拔黄山松种 好的中海拔地带常形成针阔混交林ꎮ 从实地调查
群稳定性更高(王良平和卓正大ꎬ1989ꎻ罗世家等ꎬ 可知ꎬ黄山松虽广泛分布于庐山南北山地ꎬ但大都
1999ꎻ蔡小英ꎬ2008)ꎮ 因此ꎬ在探讨黄山松种群结 冠幅较小ꎬ植株不如阔叶树种高大ꎬ不难想象在长
构和动态时ꎬ一定要充分考虑研究地的立地条件 期的森林演替过程中必然被阔叶树种所取代ꎮ 长
和群落组成ꎮ 我们的研究结论也支持以上观点ꎮ 久以来ꎬ庐山中高海拔针叶树种如日本柳杉( Cryp ̄
王良平和卓正大(1989)在对黄山松在庐山植被垂 tomeria japonica )、 日 本 扁 柏 ( Chamaecyparis
直带谱中位置讨论研究后认为ꎬ庐山生境条件较 obtusa)、黄山松等形成单优种群ꎬ林下灌木、草本
好的黄山松是森林次生演替的先锋树种ꎬ在自然 植物很难生长ꎬ对庐山水土保持、生物多样性恢
发展过程中ꎬ将被地带性植被落叶阔叶林所取代ꎬ 复、植被演替、冬季冰雪灾害防范等非常不利ꎬ因
处于向阳山顶、岭脊和陡坡等不利于植物生长的 此ꎬ鉴于黄山松更新不良现状ꎬ建议立足现有种群
高海拔山地ꎬ黄山松林才会发展成为稳定的山地 生存维持的基础上ꎬ在悬崖峭壁、山顶坡地ꎬ其他
温性针叶林ꎮ 而本文所选样地正好位于生境条件 树种难以存活的山地ꎬ可适当补植黄山松幼苗幼
较好的中山地带ꎬ我们的分析结果正好印证了王 树ꎬ人为促进黄山松种群持续稳定发展ꎻ在营养条
良平和卓正大(1989)的推测ꎮ 而经过 30 a 的自然 件较好的黄山松近自然恢复过程中ꎬ可适当补植
发展ꎬ庐山黄山松在立地条件较差的山顶、岩石 一些适宜乡土落叶阔叶树种ꎬ如小叶白辛树、短柄
上、迎风坡地等保持较好的群落ꎬ形成了一道道奇 枹、山合欢、锥栗( Castanea henryi) 等ꎬ既可平衡群
特的景观ꎮ 从庐山实地调查可知ꎬ黄山松在这种 落中黄山松小龄级个体缺失所带来的衰退趋势ꎬ
较为恶劣的生境环境下ꎬ形成优势种群ꎬ但群落中 又可以人为促进群落演替ꎬ增加群落的物种多样
黄山松幼苗、幼树依然很缺乏ꎬ黄山松的生存策略 性ꎬ以加速森林生态系统的平衡和稳定ꎮ