
Page 133 - 《广西植物》2020年第6期
P. 133
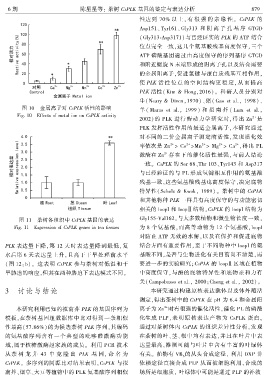
6 期 陈星星等: 茶树 CsPLK 基因的鉴定与表达分析 8 7 9
性达 到 70% 以 上ꎬ 有 很 强 的 亲 缘 性ꎮ CsPLK 的
Asp151、 Tyr161、 Gly313 和 阴 离 子 孔 基 序 GTGD
(Gly313 ̄Asp3171)与已经证实的 PLK 的 ATP 结合
位点完全一致ꎬ这几个氨基酸残基高度保守ꎬ三个
ATP 磷酸基团通过由高度保守的序列基序 GTGD
和附近螺旋 N 末端形成的阴离子孔以及结合需要
的金属阳离子ꎬ促进氢键与蛋白质残基互相作用ꎬ
使 PLK 活 性 位 点 的 空 间 结 构 更 稳 定ꎬ 从 而 提 高
PLK 活性( Kim & Hongꎬ2016)ꎮ 科研人员分别对
牛(Neary & Divenꎬ1970)、猪( Gao et al.ꎬ 1998)、
图 10 金属离子对 CsPLK 活性的影响
羊 ( Maras et al.ꎬ 1999 ) 和 拟 南 芥 ( Lum et al.ꎬ
Fig. 10 Effects of metal ion on CsPLK activity
2002)的 PLK 进行酶动力学研究时ꎬ得出 Zn 是
2+
PLK 发挥活性作用的最适金属离子ꎬ本研究通过
对不同的二价金属离子测定酶活性ꎬ发现活化效
2+ 2+ 2+ 2+ 2+
率依次是 Zn > Co >Mn > Mg > Ca ꎬ得出 PL
激酶在 Zn 存在下的催化活性最强ꎬ与前人结论
2+
一致ꎮ CsPLK 的 Ser 68、Thr 103、Tyr143 和 Asp317
与已经验证的与 PL 形成氢键相互作用的氨基酸
残基一致ꎬ这些氨基酸残基也高度保守ꎬ决定底物
特异性( Scholz & Kwokꎬ 1989)ꎮ 茶树中的 CsPLK
和其他物种 PLK 一样具有高度保守的与功能密切
相关的 loopI 和 loopⅡ结构ꎬCsPLK 的 loopI 结构为
图 11 茶树各组织中 CsPLK 基因的表达 Gly155 ̄Val162ꎬ与大多数植物和微生物长度一致ꎬ
Fig. 11 Expression of CsPLK genes in tea tissues 为 8 个氨基酸ꎬ而高等动物为 12 个氨基酸ꎬloopI
对防止 ATP 无效的水解ꎬ以及在保护和促进底物
PLK 表达量下降ꎬ第 12 天时表达量降到最低ꎬ复 结合方面有重要作用ꎬ至于不同物种中 loopI 的氨
水后第 6 天表达量上升ꎬ且高于干旱处理前水平 基酸不同ꎬ是否与生物进化有关目前尚不清楚ꎬ需
(图 12:b)ꎮ 这表明 CsPLK 参与茶树对低温和干 要进一步的实验研究ꎻCsPLK 的 loopⅡ区域在植物
旱胁迫的响应ꎬ但其在两种胁迫下表达模式不同ꎮ 中高度保守ꎬ与酶的底物特异性和底物亲和力有
关(Campobasso et al.ꎬ 2000ꎻCheng et al.ꎬ 2002)ꎮ
3 讨论与结论 本研究通过构建原核表达载体以及体外酶活
测定ꎬ得出茶树中的 CsPLK 在 pH 为 6.4 和金属阳
2+
本研究利用已知的拟南芥 PLK 的基因序列为 离子为 Zn 时有很强的催化活性ꎬ催化 PL 的磷酸
模板ꎬ在茶树基因组数据库中比对得到一条相似 化生成 PLPꎬ表明原核表达产物为 CsPLK 蛋白ꎮ
性最高(57.86%)的为候选茶树 PLK 序列ꎬ其编码 通过对茶树体内 CsPLK 的组织差异性分析ꎬ发现
的氨基酸序列含有一个典型的吡哆醛激酶功能 在茶树的叶、茎、根中均有表达ꎬ并且在叶片中表
域ꎬ属于核糖激酶超家族的成员ꎮ 利用 PCR 技术 达量最高ꎬ推测可能与叶片中含有丰富的叶绿体
从茶 树 龙 井 43 中 克 隆 出 PLK 基 因ꎬ 命 名 为 有关ꎮ 植物有 VB 的从头合成途径ꎬ 利用 DXP 非
6
CsPLKꎮ 多序列的同源比对结果表明ꎬCsPLK 与拟 依赖途径直接合成 PLP 从而被细胞利用ꎬ合成的
南芥、烟草、大豆等植物中的 PLK 氨基酸序列相似 场所是细胞质ꎬ 叶绿体中可能是通过 PLP 的补救